
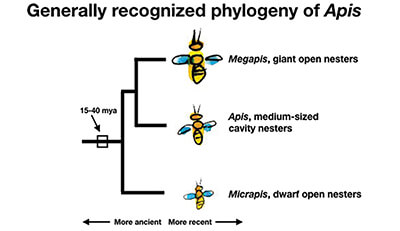
Apis. The genus to which our honey bee, Apis mellifera, belongs is so distinctive in the human imagination that its name is recycled up and down the chain of bee taxonomy. Moving first down the taxon, the genus Apis is divided into the small open-nesting species comprising sub-genus Micrapis, the large open-nesting species Megapis, and the medium-sized cavity nesting species who are granted privilege of retaining the name Apis unappended.
Moving up the chain, we have the tribe Apini, the subfamily Apinae, the family Apidae, and last of all the whole superfamily Apoidea to which all bees belong, with the curious addition – to anybody but a professional taxonomist – of a group of wasps called the Spheciformes lumped with the bees owing to shared morphology that clearly marks them as the nearest living kin of bees.
But not to despair, the “true” bees can still be pulled out of this awkward grouping and called the “Apiform apoids,” leaving one to wonder whether the clarification is worse than the problem it was supposed to solve. And then there’s the extinct genera Electrapis and Hauffapis – but at this point let’s stop, content to leave Electrapis and Hauffapis comfortably extinct and move on with our point that modern Apis, this venerable and name-giving genus we are focused on last month and next, must be important for a reason. Granted, it is the home genus of that most famous bee of all, the western honey bee, but more than that, the origin of Apis makes for a fascinating story about how and where sociality first emerged.
As my readers will eventually see, many streams of evidence are at play, but the evidence is not tidy nor consistent. The story teaches us that in the end, there is still an element of human subjectivity in judging the various streams of evidence – morphologic, geographic, genetic, behavioral, and fossil – to arrive at the most parsimonious and biologically consistent explanation for the origin and modern distribution of the genus Apis.
The Common Ancestor of Apis
But we must start our story before the emergence of Apis, variously estimated at 15-40 million years ago (mya), with the genus’s most recent common ancestor (Fig. 1). This long-lost species was a member of the so-called corbiculate bees, so named for their possession of a rear-leg pollen basket, more properly called a corbiculum (Fig. 2).
The corbiculate bees are a relatively ancient group of bees, comprising over 1,000 species, and made up of four tribes: the bumble bees (Bombini), the orchid bees (Euglossini), the stingless bees (Meliponini), and the honey bees (Apini). Lacking any proper taxonomic rank, these four tribes are nevertheless considered a “natural” grouping with a shared common ancestor from whom the corbiculum was commonly inherited.
The corbiculate bees have been the subject of considerable interest for years owing to the fact that within these four tribes we see the full gamut of social classifications, from solitary to communal (Euglossini) to primitively social (Bombini) to highly social (Meliponini and Apini). This, added to the fact that this socially diverse assemblage is nevertheless held together by one strong derived character – the corbiculum – means that the corbiculate bees constitute a tight little taxon within which we can expect to find clues to the origin of social behavior.
And with only four tribes, it should be a piece of cake to figure out their relationships and the order in which things happened, and at first this appeared to be so. Although Charles Darwin ruminated on the matter as early as 1859, it was Charles Michener in 1944, who arrived at the first, and admittedly satisfying, phylogeny based on shared behavior and morphology (Fig. 3A). This traditional phylogeny was strengthened in subsequent years by additional morphological data from adult stages, larval stages, and fossils.
Plus it has the added asset of parsimony – that philosophical bias in favor of the simplest of explanations. Parsimony, when it comes to phylogeny, means favoring a family tree that requires the fewest evolutionary changes. In Michener’s family tree, primitive sociality arose once – at the common ancestor of the bumble bees, stingless bees, and honey bees (open red circle Fig. 3A) – and advanced eusociality once (solid red circle Fig. 3A), and the more primitively social bumble bees are basal to the more socially advanced Apini and Meliponini.
A Messy Affair
But this tidy state of affairs began to unravel in the early 1990s with the advent of DNA sequencing technology that added enormously to the number of characters – in this case, shared genes – available to phylogenists for reconstructing family trees. Immediately it became apparent that trees constructed from DNA differed significantly from the traditional trees built from morphology. Moreover, the new reconstructions were not parsimonious when it came to the origins of sociality, and neither did they agree with what had been the assumed direction of change in social evolution.
This directionality, called polarity, is the vector of change over time in a character state. In the character state “sociality,” for example, we generally presume that small populations, fluid behavioral castes, and overwintering as a solitary mated female are basal states, while large populations, fixed castes, and overwintering as a colony are derived. This is the presumed polarity, or direction of change in the character state “sociality.” Moreover, it is attractive to assume that eusociality, once gained, is difficult if not impossible to reverse owing to life-history inter-dependence of queens and workers. But the new DNA trees for corbiculate bees were blowing all these assumptions to pieces.
Figures 3B-C show a phylogeny built by Sydney Cameron in 1993 based on shared mitochondrial DNA sequences. These figures highlight some of the problems this phylogeny, and others subsequently like it, caused Michener’s traditional narrative of social evolution. Cameron’s phylogeny places the highly …