
Since February of this year we have been focusing on the bee family tree and its phylogeny and biogeography that led to the western honey bee, Apis mellifera, the bee known round the world today as the honey bee. We have taken a necessarily long view, long in time that is, beginning with the corbiculate bees, that behaviorally diverse and ancient group of four bee tribes, in which at least 80 million years ago (mya) the essential building blocks of eusociality emerged.
One of those corbiculate tribes, the Apini, inherited those essential building blocks and by 15-30 mya had elaborated upon them to achieve advanced eusociality in which females multiply-mate and produce large, perennial, genetically diverse superorganismal colonies. Group selection and emergent properties synergized to create colony level functions that recapitulate similar functions in organisms.
Among the species of Apini, one of them became the common ancestor of an entire new genus, the Apis, builders of continuous regular beeswax combs, dancers of recruitment messages, and specializers in storing large quantities of nectar. It’s noteworthy that estimates for the timing of emergence of advanced eusociality in the tribe Apini (15-30 mya)1 overlap independent estimates for the emergence of Apis (30-40 mya);2-4 apparently Apis was right there at the beginning of advanced eusociality in its tribe, if not its standard-bearer.
The Outlier Apis mellifera
In February we explained that the traditional origin model places the beginning of Apis in southeast Asia where today all but one of the 9-10 recognized species are found in their natural range. That one outlier is Apis mellifera with its geographic pole set firmly in the west – Africa and western Eurasia. This geographic isolation of A. mellifera from the rest of its genus has puzzled scientists for decades and helped agitate for alternative explanations to the natural history of the genus.
In April we laid out the arguments for a European, not Asian, point of origin for Apis which, among other things, leads to a plausible explanation for the westward orientation of A. mellifera.5 Then last month we focused on the 4-5 major lineages of A. mellifera – the M, C, A, O, and possibly Y lineages – and noted their ancient migration patterns, their inter-relationships, and the names and home ranges of their member subspecies. This is where we pick up this month – peering into the natural ranges of the European lineages and considering the dynamics that led to the divergence of new subspecies within them.
The European Subspecies
I focus on European M and C subspecies for two reasons – they comprise the races of greatest interest for modern beekeeping, and their divergences present an interesting example of how subspecies divergence interacts with geography and ancient global climate.
In species like the honey bee in which genetic variation is strongly structured around geography, there are two classical explanations for the ways variants from an ancestral stock can come about.6 In our case, we consider the ancestral stock the pioneering ancestor for each of the M and C lineages, and the races within each lineage the variants.
One explanation is if the original ancestral stock was widespread and highly variable and then over time some variants go extinct, leaving distinctive races within the original range of the ancestor. A second explanation is the driver of speciation I described in February in which populations become geographically isolated from one another and, no longer capable of exchanging genes and responding to locally unique conditions, begin to follow separate evolutionary trajectories.
Ice Age Drove Lineages
Of the two, the second explanation is more amenable to constructing phylogenetic trees which are by design sensitive to “polarity” – the chronological direction of change in a character state as we discussed in March. Investigators have indeed been able to construct phylogenies of A. mellifera races. But more to the point, in the case of European A. mellifera we have ample and powerful drivers of geographic isolation and examples of its effects on honey bee subspeciation – the repeated glaciers and inter-glacial periods of the Pleistocene epoch (2.58 mya – 11,700 years ago).
The Pleistocene, or “Ice Age,” encompassed no fewer than 20 major glaciation periods, with the most extensive 5-6 occurring in the last 900,000 years.7 It is in this context that the M and C lineages advanced and settled into Europe, with expansions during the relatively warm interglacials and retreats into milder enclaves, or “refugia,” at the next cooling spell.
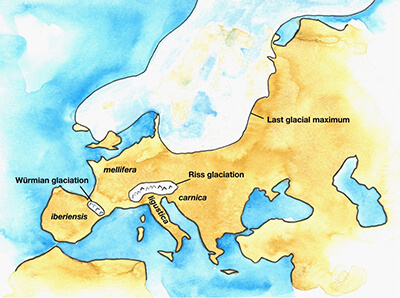
It is tempting to imagine ice age refugia as redoubts against physical obliteration from sprawling ice sheets. They were certainly that, but as Figure 1 shows, the last European glacial maximum was limited to only sections of northern Europe. There were more hazards than ice sheets, namely unfavorable weather and floral conditions. Pleistocene weather was generally drier and colder than present. There was a lot of dust. Depending on particular interglacial period, the dominant flora could range from forest ecosystems to open vegetation,8 with strong cyclic changes in availability of insect-pollinated plants.9 These repeated pulses of severe habitat change offered the ancestors of the modern M and C subspecies plenty of opportunity for divergent selective adaptation.
Moreover, glaciation could be a relatively local phenomenon. There are two examples of this, two events widely separated in place and time – the Riss glaciation over the Alps separating the Italian peninsula from mainland Europe, and the Würmian glaciation over the Pyrenees separating modern day France from Spain (Fig. 1). As we will see, these two relatively localized glaciations had outsized influence on the evolution of modern day European honey bee races.
Mitochondrial DNA estimates dating the time of divergence of A. mellifera into its constituent lineages M, C, A, O, or Y range from 1.35-0.33 million years ago (mya)10 to 0.67 mya,11 but later estimates by Wallberg et al. relying on whole-genome sequencing have edged the divergence to a more recent time around 165,000-330,000 years ago.12 Moreover, the two European lineages are surprisingly dissimilar in time and place: M split off from the ancestral A lineage an estimated 330,000 years ago and entered Europe from a western track across the strait of Gibraltar, whereas C is younger – splitting off from A about 300,000 years ago and pursuing an eastern route into Europe across the Isthmus of Suez12 (see Fig. 2D, last month’s column). Thus, M’s and C’s modern European proximity is an accident of ancient migration patterns and no product of recent kinship between the two.
There is agreement that however ancient the divergence of lineages, the radiation of subspecies within lineages is comparatively recent, specifically 38,000-13,000 years ago for M and 25,000 years ago for C,12, placing these events in the center of late-Pleistocene glacial and interglacial events.
The History of the M Lineage
Let’s look first at the elder lineage, M, which is composed of two subspecies, the northern European A. m. mellifera and the Spanish honey bee, A. m. iberiensis. The Pyrenees Mountains run in a gentle northwest to southeast vector along the borders of present-day Spain and France, as well as the microstate of Andorra. The Pyrenees have long been considered an obstacle to gene flow between the M subspecies and therefore the traditional natural range boundary between the …